Funkce formátorů a komplexů v dynamice mozkových struktur
Výzkumná zpráva č. LSS 159/03
ČVUT Praha, Fakulta dopravní
Katedra řídicí techniky a telematiky
Společná Laboratoř spolehlivosti systémů
Fakulta dopravní ČVUT Praha
Konviktská 20, 11000 Praha 1
tel. 224221721/416 (fax), 417(sekr), 418, 413 (lab)
e-mail: mirko@fd.cvut.cz
Ústav informatiky AV ČR
18207 Praha 8, Pod vodárenskou věží 2
tel. 6606/2080 , 86890639 (fax)
e-mail: mirko@uivt.cas.cz
Společná Laboratoř spolehlivosti systémů
Fakulta dopravní ČVUT Praha
Konviktská 20, 11000 Praha 1
tel. 224221721/416 (fax), 417(sekr), 418, 413 (lab)
e-mail: mirko@fd.cvut.cz
Ústav informatiky AV ČR
18207 Praha 8, Pod vodárenskou věží 2
tel. 6606/2080 , 86890639 (fax)
e-mail: mirko@uivt.cas.cz
Praha, únor 2003
Obsah:
1. Úvod
2. Analogie
3. Závěr
4. Literatura
1. Úvod
Mozek nelze pochopit jako celek, protože je "téměř nekonečně" složitý, i když z hlediska teoretického představuje konečný počet prvků, částí, drah, neuronů, synapsí, iontových kanálů, receptorů, bílkovin a molekul. Lze si však představit dílčí funkce pomocí vhodných schémat. Takové schéma může připomínat portrétní karikaturu: musí být co nejvýstižnější, ale současně co nejjednodušší. Jedním z mnoha takových schémat je model matematiků Farleye a Clarka z roku 1954.
Uvedení autoři konstruovali svůj matematický model, později realizovaný na počítači, ze čtyř základních částí: generátor signálů, komplex elementů, diskriminační jednotka a formátor čili modifikátor. Stručně vysvětlíme funkci jednotlivých částí. (obr. 1)

Obr 1.: Blokové schéma modelu Farleye a Clarka a jeho analogie s mozkovými strukturami.
Generátor signálů (thalamus) vysílá "impulzy" čili poruchové veličiny P1 a P2 do komplexu elementů (kortexu), kde je síť na počátku experimentu náhodně spojených elementů (neuronů). Během procesu učení se však již spojení mezi elementy nemění. Analytická jednotka (limbický systém) určuje stav učebního čili adaptačního procesu komplexu a informace podává jak do generátoru signálů, tak do formátoru. Modifikátor čili formátor (humoroergní modulační kmenová centra kmene mozku pro řízení vigility, spánku, nálady, epileptické a psychotické pohotovosti) řídí vstupní a výstupní prahy elementů komplexu a jejich drah. Účelem této hry je organizovat činnost komplexu a jeho odpovědi tak, aby po podráždění P1 stimulací sektoru O1 komplexu byl stimulován pouze sektor O+ a aby po podráždění P2 stimulací sektoru O2 komplexu byl stimulován čili odpovídal pouze sektor O- komplexu. To znamená, že po úspěšném "učení" umí komplex diskriminovat dva stavy (např. dvě strany, dvě písmena, dva různé podněty).
Zdůrazňujeme opět, že všechny elementy komplexu jsou na počátku pokusu mezi sebou náhodně pospojovány a jejich prahy jsou také náhodně určeny, dále i rozdělení komplexu na sektory O1, 02, 0+, O- je pouze funkční a vytváří se až během procesu učení čili adaptace na zevní (organizující) podněty.
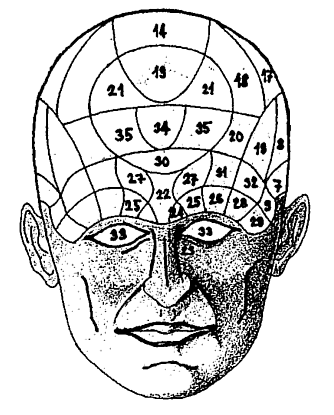
Obr 2.: Představy o lokalizaci mozkových funkcí byly různé. Marcus Marci nebo Gregor Reich se domnívali, že inteligence a paměť je umístěna do okolí komor. Před dvěma sty léty Franz Josef Gall konstruoval svou frenologickou mapu, která nepostrádá fantazii, ale je zhola vymyšlená a nezakládá se na skutečném pozorování. Ale již po šedesáti letech přichází Paul Broca se svým nálezem porušené operkulární krajiny a poruchou fatických expresivních funkcí. Gallovy mapy se uvádějí jako příklad naivity a kuriozity, zatímco Broca jakkoliv korigován, nebude nikdy naivní a bude vždy sloužit jako příklad vědeckého pozorování. Jistě by bylo zajímavé vědět, nakolik naše představy budou v budoucnu jen nepřesné nebo i kuriózně naivní. Pro ilustraci uvádíme Gallovu frenologickou mapu. Na okcipitální straně jsou "areae" 1 až 6 znamenající např. "destruktivitu, bojovnost, zamilovanost" apod., a to vše podle různých lebních protuberancií. Na frontální straně Ibi jsou to tyto "perceptivní a mentální funkce": 7 - rezervovanost či mlčenlivost, 8 - vůdcovství, 9 -konstruktivita, 10 - opatrnost, 11 - smysl pro souhlas, 12 - ocenění sama sebe, 13 - laskavost, 14 - uctivost, 15 - odhodlanost, 16 - svědomitost, 17 - naděje, 18 - smysl pro podivuhodnost, 19 - identita, 20 - veselost, 21 - napodobivost, 22 - individualita, 23 - smysl pro pořádek, 24 - smysl pro velikost, 25 - odhad váhy a odporu, 26 - smysl pro barvu, 27 - smysl pro prostor, 28 - smysl pro uspořádání, 29 - počítání, 30 - smysl pro možný výsledek, 31 - smysl pro čas, 32 - smysl pro harmonii,.33 - řeč, 34 - schopnost pro srovnání, 35 - příčinné vztahy.
Generátor signálů produkuje dva různé sledy signálů, které posílá do komplexu. Sám generátor přijímá řídící signály z diskriminační čili analytické jednotky.
Komplex se skládá ze 128 elementů, které jsou na počátku experimentu náhodně mezi sebou spojeny, ale během pokusu se jejich spojení dále již nemění. Každý element má různý počet vstupů, jejichž prahy jsou také na začátku náhodně zvoleny. Celý komplex je formálně rozdělen do 4 polí, dvou vstupních a dvou výstupních. Sledy signálů z generátoru přecházejí do vstupních sektorů (Ol a O2), a z nich jdou zcela náhodně různými cestami do obou výstupních sektorů (O + a O -).
První sekvence signálů se skládá z této posloupnosti: 001 001 001 atd. a druhá sekvence se skládá z nul: 000 000 000 atd. První sekvence se vede např. do sektoru 01 komplexu, zatímco do sektoru O2 komplexu se přivádí druhá sekvence signálů. Toto trvá určitou dobu, a po určitém čase se druhy sekvencí vymění, takže první sekvence (001 001 atd.) se vede do druhého sektoru (O2) a druhá sekvence (000 000 atd.) se vede do prvního sektoru komplexu (O1). Smysl tohoto počínání je ten, aby si model našel takovéto řešení situace: podráždí-li se sektor Ol signálovou sekvencí 001 001 atd., pak by měl být podrážděn jen sektor O +, a podráždí-li se sektor O2 signálovou sekvencí 001 001 atd., pak by měl být podrážděn jen sektor O -. První sled signálů nazývají kybernetici P l (poruchovými veličinami), druhý sled signálů P 2.
K tomuto stavu rozlišování mezi sektorem O + a O - dojde tak, že se původně náhodné spoje mezi elementy komplexu stanou organizovanějšími. To znamená, že některá spojení nabudou nižšího prahu dráždivosti, stanou se důležitějšími. Jiné spoje naopak budou mít vysoký práh podráždění, jejich spojení nebude významné a funkčně téměř zaniknou. Model hledá řešení náhodně, ale preferuje tu konstelaci spojení, která vedla ke zlepšení stavu, tedy k řešení úkolu, i když malému. Je to tzv. Hebbovo pravidlo. Tento proces představuje určitý druh učení.
Postupně se tedy v čase (t) vybudí určitý počet elementů neboli "neuronů" (N +) v sektoru O +. Nastane tak stav S l (t), nebo se vybudí ve stejném čase určitý počet elementů (N -) v sektoru O - a nastane stav S 2 (t).
Aby k tomuto organizovanému stavu došlo dosti rychle, jinými slovy aby došlo k poklesu neurčitého neorganizovaného původního počátečního stavu, tedy aby poklesla entropie systému, musí vstoupit do hry další dva členy soustavy.
Je to:
diskriminační jednotka a
formátor.
Existují však autoorganizační autonomní komplexy elementů bez "přídavných" zařízení, ale tyto elementy musí být nadány dosti velkou samostatnou pamětí, avšak i tak trvá "učení" déle než u systému podle Farleye a Clarka (Beneš, 1966, 1981).
Diskriminační čili analytická jednotka provádí analýzu stavů komplexu tím, že sleduje stavové veličiny (S), které ukazují, jak se komplex "naučil" rozlišovat dva druhy sledů signálů. Tento člen provádí jakousi diferenciaci mezi stavem S l a S 2 a to tímto způsobem: S l (t) - S 2 (t) = N + (t) - N - (t).
Jak vidíme, provádí to celkem jednoduše porovnáním počtu nabuzených elementů N ve výstupním sektoru O + a v sektoru O -. Informace o stavu organizace komplexu pak posílá do generátoru signálů a tak ovlivňuje střídání a množství sekvencí signálů do jednoho nebo druhého vstupního sektoru Ol nebo O2. Další informační kanál vede z diskriminační jednotky do formátoru.
Posledním členem autoorganizačního systému je formátor čili modifikátor. Uvedli jsme již, že tato část soustavy přijímá informace z diskriminační jednotky a po jejich zhodnocení vysílá sama signály řídícího charakteru čili akční veličiny do komplexu elementů.
Vliv formátoru na komplex má dvojí význam. Signály z formátoru jednak mění vstupní prahy čili dráždivost elementů a prahy spojů mezi elementy, a tak mění průchodnost čili vodivost drah mezi elementy, jednak z formátoru vycházejí do komplexu náhodné signály neboli šum, který má Gaussovu distribuci. Tento šum znáhodňuje deterministický proces probíhající v komplexu.
Adaptační proces systému se tak stává volnějším, a ačkoliv se to zdá paradoxní, celé učení a hledání řešení a rovnovážného stavu se tím urychluje. Formátor přijímá informace z diskriminační jednotky a srovnává počet elementů právě činných v sektorech O + a O - ve dvou časových intervalech: N (t+1) - N (t) = N + (t) - N - (t). Podle výsledků pak upravujeme prahy vstupů do elementů tak, aby bylo co nejvíce podrážděných elementů v O + při vstupu akčních veličin (001 001 ...)do Ol, anebo aby totéž platilo pro O-... O2.
Šum má velký význam pro proces automatické organizace, poukázal na to již v r. 1960 H. von Foerster. Na význam různých druhů šumů upozornila i práce Nicolise a spol. (1975). Tito autoři použili matematický model autoorganizujícího se "otevřeného nelineárního systému, který je vzdálen od termodynamické rovnováhy", a realizovali ho na počítači. Systém byl částečně podobný modelu Farleye a Clarka.
Když Nicolis ovlivňoval proces organizace systému ze zevních zdrojů intenzivním šumem, který byl amplitudově stacionární (podobal se Gaussovu šumu), organizace systému se zvětšovala, entropie klesala a stoupala redundance. Jestliže autoři použili nestacionárního "jitter" šumu, organizace se zpomalovala, popřípadě docházelo ke stoupání entropie.
My sami jsme také zkoušeli význam šumu jako nosiče regulérní proměnné a zjistili jsme, že postačují pouze 4 % harmonické regulérní proměnné "přimíchané" do Gaussovy distribuce náhodných proměnných, aby byl tento slabý, ale pravidelný signál detekován Wilksovým periodogramem (Wilks, 1967). Avšak ve Weibulově distribuci šumu, která je značně rozdílná od Gaussovy, muselo být celých 8 % harmonické pravidelné proměnné, aby mohla být periodogramem detekována (Faber a Vladyka, 1984).
V neurologické praxi to znamená, že je to především vzorec, tj. způsob výbojů hrotů, jejich sled v epileptickém ložisku, který je určující pro šíření epileptické aktivity. Není to tedy jen prostá rychlost hrotů, která definuje významnost (razanci, penetranci) epileptického fokusu (Cohen 1969).
2. Analogie
Funkční vlastnosti jednotlivých částí modelu Farleye a Clarka jsou tak zajímavé, že se sugestivně nabízejí ke srovnání s některými strukturami mozku. Generátor signálů připomíná thalamus. Tato cerebrální struktura má především dvě vlastnosti: 1. je posledním interpolačním jádrem impulzů přicházejících z kmenových jader taktilních, akustických a optických, a po zpracování vysílá tyto impulzy do kortexu, a 2. toto největší subkortikální jádro je generátorem rytmů, kterým také částečně podléhají impulzy z uvedených periferních senzorických orgánů. Thalamus má tedy i funkci hodinových pulzů (obr.3).
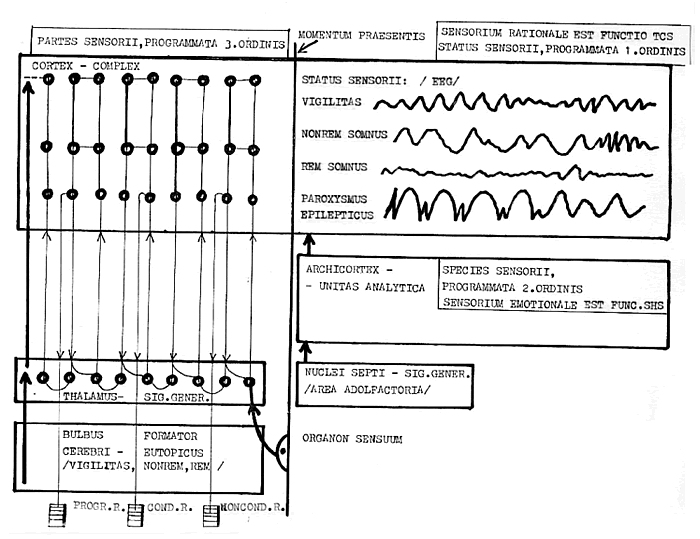
Obr 3.: Blokové schema funkčních center (funkčních bloků) v thalamu
Jednotlivá centra se v thalamu mezi sebou bohatě spojují a ještě bohatěji divergují v mnoha drahách téměř do celého kortexu.
Důkaz toho vidíme např. u evokovaných potenciálů:
již po 50 až 100 msec se šíří signál z periferních smyslů nezávisle na smyslové kvalitě (optické, akustické, taktilní signály) do celého kortexu. Tak je celý kortex informován o každé změně v okolí. Tato představa dovedena do extrému by znamenala, že každý z dvaceti miliard neuronů by byl přímo nebo nepřímo senzorickou aferentací stimulovatelný. Za fyziologických okolností tomu tak není, ale během generalizovaného epileptického výboje se stává tato představa téměř skutečností. Thalamus ovlivňuje i motoriku, o čemž svědčí bohaté spoje s nucleus ruber, s cerebellem, bazálními gangliemi a motorickým kortexem (Petrovický a Druga, 1989). Pravděpodobně zde dochází k primitivní integraci senzoricko-motorických vzruchů a tím k simplexnímu uvědomování celého těla i okolí. Někdy se thalamus nazývá bránou vědomí.
Komplex připomíná neonatální kortex, který také obsahuje iniciálně zcela "naivní, nepoučené" elementy - neurony. Teprve didaktický a pedagogický proces dítěte organizuje mozek. Vzhledem k modelu je zde řada rozdílů, např. v tom, že neurony kortexu nejsou tak zcela náhodně spojeny, ale jejich výběžky, dendrity a neurity i celé neuronální sítě rostou částečně organizovaně podle genetických pravidel již prenatálně. Vytváří se samozřejmě i makroskopická struktura mozku, hemisfér, jejich gyrifikace a základní dráhy. Vytvoří se vlastně i kortikální diferenciace na 50 Brodmannových oblastí (obr. 1, 4)

Obr 4.: Formator simposium. Liblice 1982
Mikroskopicky jsou prenatálně z části vyvinuty funkční jednotky, zvané sloupečky čili kolumny, kterých je v kortexu kočky asi 25000 a u člověka asi milion. Každá kolumna obsahuje asi 10000 neuronů a představuje skupiny neuronálních buněk ve tvaru sloupečku, který prostupuje většinou celou vrstvou kortexu. Funkčně reprezentuje tento sloupeček např. hmatové vousy (vibrissae u koček) nebo skupinu taktilních tělísek, skupinu tyčinek a čípků apod. (Petrovický a Druga, 1989). Celkový počet neuronů kortexu je asi 2x1010 .
Geneticky je tak preformována šestivrstevná struktura kortexu a některé spoje mezi kolumny, např. axony košíčkových buněk jedné kolumny zasahují do sousedních kolumn. Takto je konstruován především neokortex, který má těsné anatomické a funkční spojení s thalamem. Mezi kortexem a thalamem trvale krouží čili reverbují vzruchy, proto mluvíme o thalamokortikálnírn reverberačním okruhu. Tato reverberace je podkladem pro reaktivitu, vigilitu a bezprostřední paměť a spánek (Faber, 1992).
Jestliže se zamezí fyziologickému vstupu impulzů z oka do kortexu, pak se geneticky započatá kolumnární organizace nedokončí. U lidí od narození slepých nastane podobná situace, a dokonce se ani nevytvoří patřičná EEG aktivita mozku, tj. především základní rytmus alfa s frekvencí 8-13 Hz.
Pak jsou přítomny jiné rytmy, např. rolandický rytmus, který se u slepých lidí neovlivňuje zavíráním a otevíráním očí (Cohen, 1969). Hubel a Wiesel (1962) sešili koťatům oční víčka. Po určité době života histologicky kontrolovali stav optického kortexu a zjistili, že kolumnární struktura skutečně nebyla dokončena. Valverde (1971) ukázal, že myši chované ve tmě, mají chudě vyvinuté dendrity, trny a synapse neuronů, a naopak zvířata chovaná na světle měla neurony optické kůry na tyto organely bohaté. To znamená, že dostatečný influx impulzů do kortexu způsobí bohaté spoje mezi neurony.
Můžeme říci, že v živém organismu se "často používaný software mění na hardware" (Faber a Vladyka, 1987, Faber a Weinberger, 1988). Přeneseně můžeme říci, že opakovaná informace, např. slovo, se petrifikuje zprvu do funkčního, posléze do morfologického programu chování. Proto také při léčbě psychóz je stále nutno používat "dobré slovo" jako hlavní programovací nástroj, nestačí jen chemoterapie. Sjöström (1985) ukazuje po létech zkušeností, že schizofrenici léčení psychoterapeutiky se subjektivně cítili lépe a objektivně měli vyšší sociabilitu při podstatně menších dávkách léků než nemocní ponechaní jen na farmakoterapii. Totéž platí i pro epilepsii, kdy některé záchvaty jsou provokovány nepříznivou situací spojenou s emočním dyskomfortem, a naopak dobrou psychoterapií a anxiolytickým působením můžeme stav nemocných zlepšit (Trimble, 1985).
Podobná situace, znamenající přeměnu "software na hardware" v neuronálních sítích, se zřejmě odehrává za patologických okolností v epileptickém fokusu. Jestliže se objevuje velmi intenzivní impulzace v krysím hippokampu, dochází k zapojení excitačních neuritů zpět na vlastní neuron a tak se vytváří autopse. Tyto nové spoje představují pozitivní zpětnou vazbu, která dále aktivuje a depolarizuje neuron a vede ke zvýšené impulzaci čili "pálení". Někdy se tato "neosynaptogeneze" nazývá pučením neboli sprouting nových kolaterál i synapsí. Dále se aktivizují do té doby inaktivní synapse i iontové kanály. Takové ložisko nemusí být trvale patrné svou výbojovou aktivitou v EEG, ale má velmi snížený práh pro záchvaty a stačí jen nepatrné podněty, aby paroxysmy nastartovaly (Heinemann a Gutnick, 1993).
Dále se ve vývoji epileptického ložiska uplatní mnoho intrinsických fyziologických vlastností kortexu a hlavně hippokampálního archikortexu, kde je v raném mládí kritická perioda fyziologického vývoje, vyznačující se těmito vlastnostmi: je zde neobyčejně zvýšená pohotovost k záchvatům daná nedostatkem GABA synaptické inhibice, a naopak velmi zvýšené excitační transmise v CA3 hippokampu. Jestliže se v této kritické době objeví nějaká inzultace, jako jsou např. prolongované febrility nebo febrilní křeče, pak dojde k většímu organickému poškození neuronů, k jejich metabolickému vyčerpání během intenzivní depolarizace a vysokofrekventní impulzace (Swann, 1993).
Sumací těchto lézí vznikne velmi výkonné epileptické ložisko. Anatomicky se často jedná o tzv. mesiální temporální sklerózu. Jestliže však toto rané kritické období projde bez úhony (normální porod, žádné horečky a traumata), GABA synapse se přirozeným vývojem doplní a řada axonových kolaterál excitačního charakteru spontánně zanikne. Tato fyziologická reorganizace, spočívající ve fyziologické změně axonové arborizace a synaptoarchitektoniky, nastolí rovnováhu mezi excitačními a inhibičními mechanismy.
Avšak i tak je hippokampus slabým článkem v řetězu antiepileptických sil v mozku, protože zde zůstává poměr excitačních - inhibičních synapsí 4:1; excitační synapse přicházejí ze septálních jader, gyrus dentatus, area entorhinalis a ze Schafferových kolaterál, jediná inhibice pochází z košíčkových buněk hippokampu.
Klinické poučení je pro praxi velmi důležité, musíme zabránit vysokým teplotám a konvulzím v raném a předškolním věku, řekněme do šesti let, a to stůj co stůj. V pozdějším věku ani febrility, ani křeče a bezvědomí již neznamenají pro mozek takové nebezpečí. Jistě jsou nesprávná tvrzení, že náš mozek pracuje jen na 80 nebo 90 % apod. Obrovské množství neuronů mozku je angažováno v aktuální fázi učení. Jestliže je kognitivní (poznávací) proces a motorická odpověď na podněty již vypracována, tj. je již automatizována, pak je potřeba počtu aktivovaných neuronů podstatně nižší. Podobně i sama pasivní čili relaxovaná vigilita využívá jen málo neuronů. Jestliže se však po důležitých zevních podnětech bdělost zvýší, pak je v činnosti opět velká neuronální populace. Díky této velké redundanci počtu neuronů můžeme do kortexu "implementovat" jakýkoliv program, jak to dokazují pokusy s učením i deprivací u zvířat a nešťastné příběhy lidí, např. "vlčích dětí" nebo osud Kašpara Hausera (Veselovský, 1985). To znamená, že zdravý mozek se může naučit všemu, od schopnosti pohybovat se v džungli po možnost vědecké kariéry, tj. získat grant, vysedávat v knihovnách a v laboratořích. Mozek je univerzálně adaptabilní, má schopnost střádání všech deklarativních i nedeklarativních informací a jejich abstrakci.
Další člen automatické autoorganizační soustavy je diskriminační jednotka, kterou můžeme přirovnat k limbickému systému. Tento systém patří k fylogeneticky starým částem mozku, tj. k paleo - et archikortexu, který je u nižších obratlovců vývojově spojen s rhinencefalem, a hierarchicky je tam dominatní. U savců dominuje však neokortex, a k limbickému systému patří funkčně i některé neokortikální části, jako jsou frontoorbitální závity a gyrus cinguli.
Limbický systém (dále jen LS) dostává informace z viscerálních orgánů prostřednictvím retikulární formace a hypothalamu, a také tyto orgány ovládá cestou hypothalamo-hypofyzární neuroendokrinní soustavy a vegetativního nervstva. Kortikální část LS představuje jen 5 % plochy hemisfér, a hypothalamus má jen 0,5 % váhy mozku, a přesto tato soustava registruje a reguluje mnoho fyziologických funkcí, např. teplotu těla, osmotický tlak krve, hladinu cukru v krvi, acidobazickou rovnováhu krve a podílí se i na regulaci spánku svým jádrem nucleus suprachiasmaticus. LS přijímá i informace z neokortexu, z tzv. primárních a sekundárních asociačních polí umístěných blízko primárních kortikálních senzorických oblastí, jde tedy o nepřímý senzorický příjem optických, akustických a taktilních informací.
Septální jádra patří také k LS, ale u člověka je to area adolfactoria, která představuje "generátor hodinových pulzů", jež jsou vedeny fornixem do dalších částí, hlavně do hippokampu. Základní elektrická frekvence v tomto septohippokampálním systému je theta, u člověka asi subalfa frekvence, tj. 4-8 Hz. Základní frekvence v EEG thalamoneokortikálního okruhu je během bdění alfa, tj. 8-13 Hz. Hippokampus je vývojově archikortex, amygdala je zčásti paleokortex, jsou tu primární centra čichu, a proto drážděním těchto míst vzniká šalebné vnímání pachů, tzv. unciformní krize. Jsou-li tyto pachy velmi ošklivé, mluvíme o kakosmiích. Tyto počitky se často objevují jako aura před epileptickými záchvaty i jako halucinace u psychóz.
Celý LS má významné funkce pro tvorbu emocí, trvalých paměťových stop a významně se zde uplatňuje funkce paradoxního spánku. Zde v LS začíná stresová reakce, která postupuje do hypothalamu, hypofýzy, nadledvinek, odkud se vyplavují hormony působící spolu s vegetativními nervy "pohotovostní", sympatikotonickou reakci těla. Při tomto procesu stoupá puls, krevní tlak, svalový tonus, rozšiřují se štěrbiny oční a pupily a současně se objevuje většinou negativní emoční reakce v podobě zlosti a tendence k agresi, nebo naopak k defenzivě. V hippokampu je umístěn jakýsi časový registr, který třídí paměťové stopy podle toho "kdy a kde se odehrály", hlavně to platí pro deklarativní paměť. Tento paměťový děj je vždy závislý na afektu. Zapamatování, stejně jako veškerá mentální činnost, je vždy doprovázena nějakou emocí. Čím je emoce silnější (a mírně pozitivní), tím je zapamatování dokonalejší. K učení vždy přistupujeme s nějakým motivem, zaujímáme k němu určitý postoj. LS tedy spojuje racionální a emocionální chování a vytváří tak strukturu osobnosti.
Motivy lze různě třídit, např. rozeznáváme biologický a sociální původ motivů. Primární čili biologické motivy jsou např. instinkty. Jsou to vlastně složité vrozené nepodmíněné reflexy. Např. ptáci na jaře stavějí hnízda, rozmnožují se, mají rodičovské pudy, a naopak na podzim se houfují a odlétají. Podnětem je zde přibývání nebo ubývání světla. Provedeme-li léze v hypothalamu, septu nebo hippokampu, toto chování se poruší. Anatomická dráha takového a podobného mechanismu chování vede z retiny do hypothalamu, odkud se neurokrinním způsobem ovlivňuje sekrece hormonů neurohypofýzy a releasing hormony, vylučovanými do portálního oběhu, se stimuluje sekrece hormonů adenohypofýzy. U lidí podobným motivům ze sféry psychosexuální říkáme impulsy.
(Slovo impuls má více významů, jednak je používáno pro označení velmi krátkého signálu, dále se používá pro uvedený druh motivů, též pro elektrochemický děj čili vzruch, který přenáší podráždění z jednoho neuronu na druhý, a konečně toto slovo znamená náhlé, prudké patické chování, např. suicidální úmysl a jeho realizaci u psychotiků.)
Impulsy v pojetí této zprávy představují endogenní tendence k chování, probouzejí motivy a podílejí se na vrozené části osobnosti. Lidé jsou geneticky determinováni jako sangvinici, cholerici, flegmatici a melancholici nebo mají sklon k rozumovému či uměleckému myšlení apod. (Madsen, 1972). Různí psychologové a jejich školy vytvořili důmyslné kvalifikační systémy osobnosti. Např. Eysenck rozlišuje v osobnosti škálu neuroticismu, kde polární extrémy jsou supresoři a senzitoři, a v kolmé rovině na předešlé vlastnosti klade stupně "sociability", kde jsou polární extrémy extroverti a introverti.
Vedle impulsů rozeznáváme incentivy, které představují část motivů navozených určitým druhem výchovy, především v raném mladí, většinou v rodinném prostředí a částečně ve škole. Incentivy usměrňují motivované chování. V ideálním případě jdou impulsy a incentivy ruku v ruce. Např. dítě má hudební nadání, a alespoň jeden z rodičů je hudebník. Dítě vyrůstá v harmonickém vztahu svých impulsů a incentiv, tj. učí se hudbě, baví ho to, má úspěch a je chváleno.
Samozřejmě existuje mnoho přechodů od ideálního vztahu impulsů a incentiv až k pesimálnímu vztahu, kdy je např. pubescent impulzivně (instinktivně) hypersexuální, ale vychováván je v rodině nábožensky nebo puritánsky založené. Vyvíjí se pak endogenní frustrace čili střetávání dvou různých motivů, impulsy jsou hypersexualita, incentivy jsou přísné mravní příkazy, což lze nazvat konflikt.
Další vývoj takového mladého člověka závisí na jeho dalších povahových vlastnostech. Nedojde-li k naplnění motivovaného chování, dochází k frustraci či úplné deprivaci instinktivní či obecněji cílové potřeby. Jedinec na to pak reaguje podle svého charakteru depresí, regresí (např. infantilismem) nebo agresí. Konflikt bývá doprovázen objektivně společenskými dramatickými scénami a subjektivně nepříjemnými emocemi, často i s příznaky stresu. Uvedený proces je klasická cesta k neuróze, vzniklá na základě konfliktu motivů. Celý děj může probíhat nevědomě. Dobrý psychoanalytik tyto inaparentní konflikty může poznat a pomoci nemocnému tím, že mu latentní psychotraumata pomůže odhalit. Spolu se pak mohou pokusit evokovat traumatizující situaci při plném uvědomění si všech okolností, zbavit ji důležitosti, bagatelizovat ji a ukázat nemocnému její "neškodnost". Tak lze změnit náhled pacienta na vlastní potíže a zbavit ho příčiny k náhlým nebo chronickým úzkostem či somatickým příznakům, jako jsou např. palpitace, dyspepsie, kolísavá hypertenze, cefalea, a snad zlepšit i somatický patický stav, jako je např. gastrointestinální ulcerace apod. Tyto vegetativní a viscerální příznaky totiž často doprovázejí tzv. neurotické stavy.
Tato fakta jsme museli uvést dříve, než se pokusíme v kapitole o epileptogenezi identifikovat mechanismy koincidence epileptogenních a psychogenních traumat, je-li přítomna organická léze a epileptický fokus v limbickém systému. Vzniklý syndrom označujeme jako epileptóza. LS v mozku, stejně jako diskriminační jednotka v modelu, trvale monitoruje stav viscerální, somatický a neokortikální, tj. aktuální obraz o vnitřním prostředí organismu a o postavení živočicha v přírodě a člověka ve společnosti. LS vše vzájemně integruje a vytváří reflexně stav okamžité psychosomatické situace, kterou nazýváme forií nebo afektivitou. Tato může být neutrální nebo pozitivní (radost) nebo negativní (smutek, zlost), a je vždy doprovázena nějakou vegetativní reakcí a často i sekrecí hormonů.
LS reguluje stavy organismu i forii pomocí vegetativního nervstva i endokrinie. Gallagher (1987) sledoval epileptiky pomocí zanořených elektrod a zjistil, že epileptické výboje v hippokampu kratší než 10 vteřin inhibovaly hypothalamo-hypofyzární systém a tlumily sekreci ACTH. Výboje delší než 10 vteřin nebo sám psychomotorický paroxysmus naopak zvyšovaly sekreci ACTH i prolaktinu. Tento poznatek modeluje vliv LS na stres: krátké výboje představují jakousi hyperfunkci hippokampu, a tedy jeho tlumivý vliv na hypothalamus, a naopak delší výboje reprezentují již hippokampální hypofunkci, a proto je zřejmě hypothalamus deliberován a je hyperfunkční.
LS systém má asi hlavní podíl na fenoménu, který můžeme nazvat "uvědomování si skutečnosti" (Faber a Vladyka, 1987). Jak jsme již naznačili, LS percipuje interoceptivní - viscerální, somatické - taktilní a proprioceptivní a exteroceptivní stimuly, integruje je, a tak asi vzniká emotivní náboj a somatizace prožívaného. Můžeme shrnout: kmenové a limbické struktury jsou místa, kde se "somatizují" psychické děje převodem do somatické a viscerovegetativní oblasti, a naopak somatické děje se transformují do psychiky s emočním egocentrickým nábojem. Během paradoxního spánku se tyto struktury velmi aktivují, proto máme také při snech pocit, že jsou naše prožitky reálné.
Za patologických okolností vzniká v LS iritační aktivita, která vede k epileptickým psychomotorickým záchvatům nebo k psychotiformním stavům. V pokusech na opicích bylo zjištěno, že model psychózy po lysergové kyselině (LSD) se nezmění po frontální lobektomii, ale po temporální lobektomii jako by ani psychóza neprobíhala, většina příznaků po LSD chybí. U epileptického záchvatu temporálního laloku nejde o bezvědomí, ale o ztrátu funkce paměti, zmizí reálný kontakt s okolím a nastartuje nějaký neadekvátní motorický automatismus nebo konfúzní chování. Pomocí zanořených elektrod jsme se o tom opakovaně přesvědčili. V amygdale a hippokampu byla nalezena intenzivní epileptická aktivita během psychomotorického záchvatu, zatímco ve skalpových, tedy neokortikálních elektrodách byl často záznam téměř normální nebo jen lehce abnormální.
LS má kromě uvedených funkcí (motivace, afektivita, pocit reality) zásadní vliv na paměť a podílí se i na věcném myšlení. Správná činnost fyziologických funkcí limbických je asi udržována periodickým opakováním paradoxního spánku. Deprivace paradoxního spánku poněkud připomíná Klüverův-Bucyho syndrom: hyperfagie, hypersexualita, hypomnézie, hypermetamorfismus. Poslední symptom je určitý druh "roztěkanosti", neschopnosti soutředit se na jednu věc. Epileptické výboje v LS způsobují psychomotorické záchvaty a jim podobné simplexní i komplexní parciální záchvaty.
Funkce thalamoneokortikálního systému jsou vědomí, vigilita, synchronní spánek, reaktivita, abstraktní myšlení.
Za patologických okolností při epileptických výbojích se objevuje bezvědomí; jde o centrencefalické záchvaty, jako je absence (petit mal), a jestliže se patologické impulzy iradiují do kmenových struktur, dochází k tonicko-klonickým generalizovaným křečím (grand mal). Poslední část systému autoorganizace je formátor, který připomíná činnost kmenových modulačních humoroergních center. Mozek je podstatně složitější "autoorganizační" systém než uvedený model, proto má těchto formátorů více. Tyto formátory slouží k udržování vigility, spánku, nálady a nepřímo i učení a paměti.
Ukládání informací do paměťového média je v neživém stroji podstatně jednodušší než v neuronech. Stroj "procesuje", tj. pracuje s daty, jen tehdy, jsou-li v logicko-aritmetické jednotce, ale mozek s nimi pracuje téměř stále a nelze dobře odlišit takovou procesující jednotku od paměťového média, je to neustále pracující živá neuronová síť.
Mohli bychom zjednodušeně říci, že paměť je v každém neuronu a každý neuron je malým hybridním procesorem.
Toto je velmi stará myšlenka, vyslovil ji podobně již Jan Marek, náš slavný, ale zcela zapomenutý rodák před třemi sty lety. Takto je doslova tato idea vyřčena v knize "Liturgia mentis: .... ita ut in quocunque illius puncto sit tota, et quodlibet punctum speciei in toto". Volně přeloženo: autor zde říká, že (paměť) je ve kterémkoliv bodě celá a kterýkoliv bod (paměťové stopy nebo řekněme viděného obrazu) je v celé (paměti) (Servít 1989). Je to tedy holografický princip zobrazování, který postuloval pro práci mozkové kůry až v současnosti Barret (1970).
Jeden z formátorů, který byl objeven jako první, je ARAS (ascendentní retikulární aktivační systém). Je uložen v pontu a mesencefalu a slouží mimo jiné k udržování vigility a svalového tonu. Fréderic Bremer v pokusech na kočkách před 2. světovou válkou chirurgicky transsektoval interkolikulárně mesencefalon, a zvíře bylo uvedeno do bezvědomí. Experimenty postoupily až po válce, kdy Moruzzi a Magoun zjistili pomocí lézí kmene, že se nejedná o celý mesencefalon, který se na řízení vigility podílí, ale jen o jeho centrální část, tzv. tegmentum (Moruzzi, 1949, 1964). Acetylcholinergní neurony tegmenta svými axony stimulují thalamus a nepřímo kortex. Vhodnou frekvencí thalamokortikální reverberace je udržován živočich bdělý. Lze říci, že ARAS je formátorem pro vigilitu. (obr. 5).
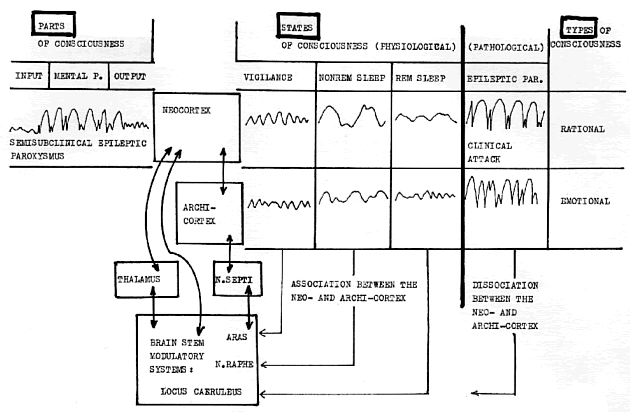
Obr. 5.(NNW 1991 vol. 1. no. 1):Attempted interpretation of the complex notion consciousnes (sensorium), according to electrophysiological findings. Parts of consciousness can he classified as input: sensory function (seeing, hearing), mental processing: (MENTAL P.) (gnostic function, abstract thinking), output: psychomotor activity (behaviour, speech). States of consciousness are vigilance. NONREM sleep and REM sleep, pathological stales are e.g. epileptic-paroxysms or psychotic raptures. Types (or kindes) of consciousness are rational and emotional. Left above: EEG curve of semisubclinical epileptic paroxysm, the higher mental activity disappears, but simple reactivity to external stimulation remains. Right above:
formators of synchronnous sleep.
Po určité předehře synchronního spánku, který trvá v prvním cyklu NONREM-REM asi 2 hodiny, nastupuje tzv. paradoxní spánek čili REM, "rapid eye movement", tedy spánek s rychlými pohyby očí, které jsou pro tento spánek (a pro bdělost i při zavřených očích) typické. Uvedený spánek trvá asi 10-15 minut a přechází opět do NONREM čili synchronního spánku. Takto se cykly NONREM-REM opakují asi 4-6 krát za noc a trvání REMu se v každém cyklu stává delším a trvání NONREMu kratším. Centra kmene, která startují a řídí REM spánek, jsou noradrenergní locus caeruleus a acetylcholinergní nucleus gigantocelularis v pontu. Rozsah divergence axonů těchto jader je obrovský, ovlivňují kmenová centra, thalamus, bazální ganglia, limbický systém i kortex. Uvedená jádra jsou formátory pro paradoxní spánek. Celkové množství paradoxního spánku je u dospělého člověka asi 20-25 %.
Všechna popsaná centra (formátory) ovlivňují mozkové i somatické funkce nejen přímo svými centrifugálně jdoucími axony, ale i humorálně, protože regulují hypothalamo-hypofyzární sekreci. Tak např. v hlubokém NONREM spánku se vylučuje somatotrofní hormon, ve druhé polovině noci se vylučují ACTH a kortikoidy, v REMu se vylučuje prolaktin, testosteron, endorfiny apod. Každý stav, tj. vigilita, synchronní a paradoxní spánek, se vyznačuje svou specifickou makro- i mikro- EEG aktivitou, humorální a hormonální sekrecí i psychickou činností. Tak v klidné bdělosti a v NONREMu pracují neurony relativně pomaleji a v jiném vzorci výbojů, zatímco v REM spánku je frekvence neuronálních vzruchů rychlejší, a tyto pálí ve skupinách čili vytvářejí "grouping" podobně jako v aktivní bdělosti. REM je doprovázen živými sny s emočním podtextem. Podle amerických statistik prožívají i zdravé osoby ve snu ze dvou třetin negativní emoce. V REMu jako v bdělosti jsou aktivní nejen senzorické systémy, ale i asociační a všechny motorické oblasti kortexu a dále tj. nucleus ruber, cerebellum, bazální ganglia, ba i pyramidová dráha. Motorické dráhy jsou však na úrovni periferních motoneuronů kmene a míchy blokovány, asi presynaptickou inhibicí. Tak je vyloučeno, že to, co se nám zdá, nemůžeme skutečně pohybem realizovat. Takové mechanismy jsou vyvinuty asi u všech savců.
Jouvet a Delorme (1965) však provedli lézí v distální části locus caeruleus. Takto preparované pokusné zvíře, byla to kočka, předváděla svými pohyby v paradoxním spánku to, co se jí zdálo. Zřejmě vše, co prožíváme ve snech, není jen na úrovni senzoricko-imaginativní, ale i motorické a psychomotorické. To jest, zdá-li se nám o plavání či běhání, pak skutečně v mozku vytváříme specifické a konkrétní pohybové neuronální vzorce výbojů pro plavání a běhání, a jen blokáda motorických drah v poslední instanci ve kmeni a míše, v motoneuronech periferní motorické dráhy, nám nedovolí, abychom tyto pohyby dovedli do konce, tj. uskutečnili je.
"Jouvetova kočka" dovoluje vysvětlit některé syndromy z lidské patologie spánku, např. somnambulismus nebo pavor nocturnus, a snad i některé druhy nočních psychomotorických epilepsií. V těchto případech předpokládáme nedostatečnou presynaptickou inhibici periferních motoneuronů, která fyziologicky nastává během paradoxního spánku. Zřejmě je realizována retikulospinální drahou. Za uvedených patologických okolností jde tedy o deliberační fenomén, způsobený buď přílišnou intenzitou REMu nebo nedostatečnou myelo- či synaptoarchitektonikou retikulospinálních drah, vedoucích z locus caeruleus do kmene a míchy.
Složitá organizace spánku savců má asi několik funkcí. Jednak je to jistě restaurační somatický proces, jednak asi slouží ke kontrole získaných informací a k ukládání individuálně důležitých informací do paměti. Bylo zjištěno, že během NONREM spánku se v neuronech zvyšuje množství RNA a během REM spánku stoupá množství proteinů. Zamezíme-li pokusné osobě spát spánkem, hlavně REMem, pak se velmi zhorší zapamatování nových informací a klesne inkorporace aminokyselin do proteinů.
Epileptický fokus působí pravděpodobně také jako formátor. Všimněme si vlastností epileptického ložiska. Jak jsme uvedli již výše, vývoj epilepsie se zakládá na třech kategoriích: vznik ložiska, zvýšená záchvatová pohotovost a přítomnost vyvolávajícího epileptického podnětu. Po biologické stránce je funkce i struktura ložiska dosti dobře definovaná. Je zajímavé, že se podle etiologie podstatně neliší, tj. má podobné vlastnosti, ať vznikne na podkladě asfyxie, traumatu nebo zánětu (Benešová a spol., 1980). Z etiologického hlediska jsou velmi nebezpečné a časté perinatální asfyxie, později vleklé horečky nebo již probíhající paroxysmy s prolongovaným trváním. (obr. 6).
V epileptickém ložisku degeneruje glie, neuron se ocitá v havarijním stavu, zanikají axosomatické inhibiční synapse a dendritické modulační synapse (Fischer a spol, 1968). Z experimentálního hlediska máme k dispozici řadu modelů ložisek, jeden z nich vzniká elektrostimulací. Během jemné, ložiskové, těsně nadprahové, pravidelné, ale sporadické elektrické stimulace zvané kindling, stoupá výkonnost receptorů, především excitačních, např. NMDA (N-metyl, D-aspartátových). Bylo zjištěno, že v mozku epileptiků je naopak málo inhibičních transmiterů -serotoninu i GABA, což lze někdy potvrdit nálezem nízké hladiny těchto látek v likvoru. Tyrosin jako další inhibiční transmiter má dokonce snížené vylučování v moči epileptiků.
U geneticky podmíněných epileptických myší je vrozený deficit GABA-benzodiazepinových receptorů a aktivita ATPázy je snížena. Rovnováha mezi natriem a kaliem v extra- a intracelulárním prostoru je udržována právě pomocí ATPázy v neuronálních a gliálních membránách. V záchvatovém ložisku se kalium hromadí v intracelulárním prostoru, což vede k depolarizaci. Látky, jako oubain, které specificky blokují ATPázu, vyvolávají paroxysmus. Dieta chudá na pyridoxin (vitamin B6) může také vést ke zvýšení epileptické pohotovosti, protože tento vitamin je kofaktorem mnoha enzymů, mimo jiné glutamát-dekarboxylázy, která kromě dalších enzymů syntetizuje GABA kyselinu.
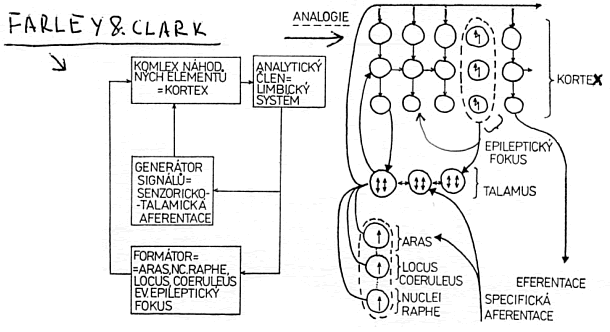
Obr. 6.(Čas. Lék. čes., 120, 1981, č. 44): Vlevo autoorganizační kybernetický systém, jak jej publikoval Beneš; vpravo analogické přiřazení mozkových struktur k tomuto autoorganizačnímu modelu. Velmi udivující je trvalá aktivita epileptického ložiska, která ve spánku dále stoupá, přesto však nedochází ke stále se opakujícímu nebo stále trvajícímu epileptickému záchvatu.
Jak jsme již uvedli, v epileptických ložiscích trpí i glie. Nepřímo pak je touto insuficientní glií postižen i neuron. Normální gliové buňky se podílejí na metabolismu neuronů, např. dotýkají se svými výběžky jak kapilár, tak i stěn neuronů, a zprostředkují tak kontakt neuronů s krevním oběhem. V epileptickém fokusu tato výkonná glie zaniká, proliferují zde tzv. fibrózní astrocyty a vzniká glióza čili gliová jizva. Obvykle se tak děje v subpiálních čili marginálních oblastech. V amygdale nebo hippokampu je pak popisována tato glióza jako mesiální temporální skleróza, a v cerebellu jako lobulární skleróza.
Neuron není v ložisku dostatečně inhibován, sám je navíc metabolicky poškozen, je depolarizován, deaferentován ztrátou apikálních dendritických synapsí, dendritických trnů a axosomatických synapsí. To znamená, že není již zapojen do neuronální sítě svými aferenty, ale sám svým neuritem může ovlivňovat další neurony. Takto depolarizován a deliberován má tendenci vydávat vzruchy o neobvykle vysoké frekvenci. Také jsme již uvedli, že někdy vyrůstají excitační kolaterály z vlastních axonů zpět na somatodendritickou membránu, vytváří se tzv. sprouting čili autopse. Takto vzniklá pozitivní vazba vede k dalšímu zesílení impulzace. Hippokampus savčích mláďat, a tedy i člověka oplývá fyziologicky množstvím excitačních a nedostatkem inhibičních GABAergních synapsí.
Po vysokých horečkách jakékoliv etiologie se tato situace ještě zhorší. Totéž nastane po opakovaných delších paroxysmech, snad hypoxií nebo "metabolickým" vyčerpáním nervové tkáně. Když tato ložisková destrukce nastane v hippokampu, který má sám o sobě velmi nízký záchvatový práh, pak je epileptogeneze velmi rychlá a postihne neblaze i paměť a psychiku. Hippokampus má i de norma 4krát více excitačních než inhibičních synapsí. Epileptické ložisko se však může vyvinout v kterékoliv části starého i nového kortexu, avšak v archikortexu se projevuje z uvedených důvodů maligněji.
K vývoji fokusu přispívá nejen akutní trauma či asfyxie, ale i chronické změny, jako je působení např. železa uvolněného z vlastních erytrocytů. Kromě vzácných inertních kovů (zlato, platina) mohou snad všechny kovy působit epileptogenně. Snad je to katalytický vliv na metabolismus vody. Pod vlivem železa vzniká množství peroxidu vodíku, který dále poškozuje neurony. Tkáňové peroxidázy a katalázy již nestačí masivně vznikající peroxid vodíku rozkládat (Willmore a Rubin, 1981). U jiných kovů, např. u olova či kobaltu, se navíc uplatňují i toxické vlivy.
Dále vznikají nad ložiskem fibrózní meningocerebrální jizvy, které mohou dráždit či zhoršovat vaskularizaci ložiska a vést k další hypoxii. Na ischemii jsou citlivé 3. a 4. vrstvy neokortexu, hippokampus, thalamus a bazální ganglia. Tyto struktury mají relativně vyšší obsah GABA, především v malých inhibičních interneuronech, které jsou citlivější na hypoxii než velké motorické neurony. Množství GABA se po ischemii sníží. I v temporálních epileptických fokusech lidí bylo nalezeno snížené množství dekarboxylázy glutamové kyseliny, což je ferment, který je zapojen do procesu syntetizujícího GABA kyselinu.
Jako biologické modely dobře slouží experimenty na zvířatech, protože pálení neuronů v epileptickém ložisku, např. po přiložení aluminiového gelu na kortex zvířete, je téměř identické s lidskými fokusy, jejichž elektrická aktivita je snímána peroperačně (Ward, 1969). Známe dobře rychlost a strukturu jednotlivých výbojů epileptických neuronů. Rychlost dosahuje 200 až 900 impulzů za vteřinu, zatímco normální frekvence je několik impulzů až desítek impulzů za vteřinu. Struktura výbojů neuronů v epileptickém ložisku je buď jednoduchá, kdy neuron pálí stále stejně vysokou frekvencí "tonicky", nebo složitá, kdy pálí "klonicky" v salvách nebo i v jiných vzorcích o různé sekvenci impulzů. Normální výboje normálních neuronů jsou nepravidelné a relativně pomalé (Creutzfeldt a Houchin, 1974, Machek, 1965, Crandall a spol., 1972, Lange a spol., 1983).
Calvin (1972) měřil mnoho lidských i experimentálních epileptických neuronů a průběhy těchto salv pak modeloval na počítači. Zjistil překvapující věc, že postačuje pouze l % velmi rychlých impulzů dopadajících na zdravý neuron, aby se i tento stal prostorovou a časovou sumací impulzů epileptickým, a také začal pálit vysokofrekvenčně. Toto zjištění svědčí o fyziologické "náchylnosti" mozku k epileptickým záchvatům a o důležitosti inhibičních neuronů. 30 % synapsí v mozku je GABAergních. Při velkém množství neurotransmiterů je to naprostá většina této aminokyseliny mezi mediátory. Jak jsme uvedli ve stati o antiepileptikách, postačuje často blokovat jen třetinu těchto inhibičních GABAergních synapsí, aby došlo k epileptickému záchvatu.
Kromě GABA má inhibiční efekt i glycin a taurin, somatostatin a serotonin, do jisté míry i noradrenalin a někdy i dopamin. Prekurzor serotoninu 5-hydroxytryptofan je účinný proti křečím po cerebrální asfyxii. GABA spolu se serotoninem účinkují nejen na úrovni kortexu, ale i v bazálních gangliích a ve kmeni. Serotoninergní systém reguluje synchronní spánek, vyvažuje emotivitu, je to mediátor euforie. Působí jako antagonista proti všem ostatním hlavním humoroergním systémům a zpomaluje jejich aktivitu. Jestliže nastane hyposerotoninergie, pak se objeví jakési překotné, zbrklé a nepřesné reagování zvířat v behaviorálních experimentech. Stav se trochu podobá Klüverovu-Bucyho syndromu. Průmyslově se vyráběl 5-hydroxytryptofan pod značkou Levothym, indikace byly deprese a insomnie. Epileptici někdy mívají sníženou hladinu serotoninu a 5-hydroxyindoloctové kyseliny v moku.
Sloviter (1987) popisuje velké snížení somatostatinergních neuronů v hippokampu potkanů po intenzivnější stimulaci vedoucí k epileptickým záchvatům. Také téměř zmizely mechové buňky (mossy cells), které jsou v hilu gyri dentati. Košíčkové GABAergní buňky byly v tomto experimentu kupodivu málo poškozeny. Autor se domnívá, že jde spíše o poruchu těchto hilárních interneuronů, které pak nemohou aktivovat košíčkové buňky, a ty potom nevykonávají svůj velký inhibiční vliv na pyramidové neurony hippokampu.
Jak jsme již uvedli, poškozené neurony v epileptickém ložisku jsou deaferentovány a deliberovány a nalézají se v parabiotickém stavu, proto patologicky pálí, tj. vysílají velmi rychle své impulzy svými zachovalými neurity k dalším neuronům. Takových vysokofrekvenčních neuronů je v ohnisku asi 10 %. 40 % neuronů je fakultativně epilepticky pálících, tj. vybíjejí se občas normálně čili pomalu a dle potřeby neuronální sítě a dle stimulace, občas však pálí patologicky jako uvedených 10 % obligatorně pálících epileptických neuronů. Zbývajících 50 % neuronů pálí v ložisku fyziologicky.
Důležitá je funkce oněch 40 % neuronů, jejichž aktuální stav určuje, zda dojde k nadkritickému množství epilepticky vybíjejících neuronů. Jestliže k takovému stavu dojde, pak nastane proces iradiace epileptické aktivity do okolí, vznikají sekundární ložiska a během dalšího působení na thalamus dochází ke generalizaci paroxysmálních výbojů na celý kortex a může se objevit klinicky manifestní epileptický záchvat.
Skupina nenormálních neuronů v epileptickém fokusu představuje jakéhosi "vetřelce", který se vymanil z integrity kortexu a stal se "samozvaným" donátorem patologických rytmů. Fokus zaplavuje asociační a komisurální vlákna intenzivní abnormální impulzací a vytváří tak sekundární a terciární fokusy, aktivuje a "učí" thalamus také abnormálnímu neuronálnímu pálení. Když dojde k určité impulzační rezonanci mezi epileptickým fokusem a thalamem, objeví se subklinická nebo i klinická manifestace elektrických epileptických výbojů.
Fokální aktivita zasahuje rušivě i do kmenové regulace. Ve kmeni pak dochází k poruchám sekrece hormonů hypothalamo-hypofyzárního systému, k dysregulaci imunomodulačních mechanismů a spánkových center. Objevuje se deformace spánku (nedostatek REMu u epileptiků, nedostatek NONREMu u schizofreniků) a paměti. Někdy se zhoršuje nálada a objevují se i úzkosti, a to nejen ze sekundárních příčin, protože si nemocný uvědomuje svou chorobu, ale i z příčin primárních, tj. z neblahého působení ložiska na humoroergní systémy.
Proto nazýváme epileptický fokus aberantním (zcestným) a ektopickým (mimo kmen lokalizovaným) formátorem. Tento abnormální formátor soupeří o vliv na thalamokortikální a septohippokampový okruh spolu s fyziologickými formátory. Jakmile fyziologické formátory prohrají a neudrží svůj vliv, dochází k epileptickému záchvatu nebo psychotické krizi.
Modelovali jsme spolu s dr. Novákem v České akademii věd funkci epileptického ložiska na okolí pomocí umělých neuronových sítí, které měly v každé ze dvou vrstev po 5, 10 nebo 50 "neuronech". Funkce sítě, která je schopna se něčemu naučit, např. rozeznávat určité tvary křivek, se může dále zlepšit, jestliže se v první vstupní vrstvě poněkud sníží vstupní práh podráždění v ohraničené skupině sousedních "neuronů". Když se vstupní práh podráždění ještě dále sníží, rozeznávací funkce se nezlepší, ale naopak zhorší. Toto je simulace fokální léze, která, jak vidíme, může vést nejen ke zhoršení, ale i ke zlepšení pracovního výkonu. Jestliže je takto porušena více než pětina vstupních elementů, pak je síť již velmi poškozená. Svědčí to o tom, že malé ložiskové poškození nemusí znamenat pro složité systémy žádnou škodu. Poškození mediálních elementů je pro funkci sítě horší než laterálních, což připomíná klinickou zkušenost těžkých příznaků při lezích primárních projekčních areí a nevelké příznaky i při rozsáhlých lezích v asociačních oblastech.
Změníme-li prahy různých nesousedících neuronů, tj. vytvoříme-li model difúzní léze, pak je porucha funkce výraznější i při stejném počtu poškozených "neuronů" jako v ložiskové lézi. Tento stav modeluje těžké změny inteligence při difúzních atrofiích mozku.
Obecně lze říci, že velmi složité systémy v živé i neživé přírodě, např. systém počasí na zeměkouli nebo život v moři, jsou velmi rezistentní proti poruchám. Mohou být zničeny rozsáhlé části systému, aniž by došlo k závažným poruchám. Avšak tyto systémy mají kritické body, kde i malá porucha vede ke katastrofálním následkům. V mozku je to např. ARAS kmene, kde malá léze způsobí velké změny vědomí nebo jádra pro spánek, jejichž porucha způsobí deformaci spánku až insomnii, nebo malé léze v hippokampu, jejichž přítomnost vede okamžitě k amnézii apod. Podobně i epileptický fokus působí nejen podle své intenzity (velmi rychle pálící neurony jsou nebezpečnější), ale i podle lokalizace. Jak jsme již uvedli, frontálněji umístěná ložiska jsou více epileptogenní než ložiska umístěná parietálně či okcipitálně. Temporálně lokalizovaná ložiska vedou nejen k těžkým formám epilepsie, ale často i k devastaci psychiky, paměti, emoční rovnováhy a osobnosti.
Klinická pozorování a výsledky modelování naznačují i opačnou možnost. S jistou licencí lze vyslovit domněnku o "benefitním" působení neintenzivních epileptických ložisek v neepileptogenních oblastech parietálních (a posteriorních temporálních ?). Zde lokalizované fokusy snad mohou vést i ke stimulaci mozku směrem k pozitivním výsledkům, jako je zlepšená mentace, zvýšená kreativita apod.
Neurofysiologická aktivita neuronálních impulsů je však především základem fysiologické percepce a učení. Kupř. děti v l. třídě se učí kreslit různé čáry a z nich sestavovat a psát písmena. Je to stále pokračující proces od kreslení a psaní po věcné a posléze abstraktní myšlení. Té počáteční situaci se dá říkat soubor počátečních podmínek čili basén atrakcí. Celá (Obr. 7) transformace od čar, písmen, slov viděných (grafém) nebo slyšených (morfém) až po psychický pojem (sémém) se fakticky uskutečňuje přeměněním sensorických podnětů na sekvence neuronálních impulsů (SNI). Tyto impulsy jsou fysikálními nosiči čili signály nižší i vyšší nervové činnosti - informace. Smyslový podnět i mentace jsou zakódovány v SNI. Již jsme definovali perceptogram jako zakódované SNI, které jdou od smyslových orgánů do thalamu. Mezi thalamem a kortexem již není přetržitá impulsace jako v perceptogramu, ale trvalá reverberace impulsů, kterou nazýváme ideogramem. V ideogramu dostává kódování představ obecnější charakter a uskutečňuje se zde vlastní kognice. Ideogram je podkladem mentačních procesů. Perceptogram transformovaný na ideogram v thalamokortikálnírn reverberačním systému je vyjádřen určitou funkcí, kupř. funkcí atraktoru. Tato transformace znamená pro nervovou sít určitý návod, jak zacházet se zevními stimuly, jak je poznávat. Je to kognitivní funkce a lze ji nazvat algoritmem. (obr. 8)
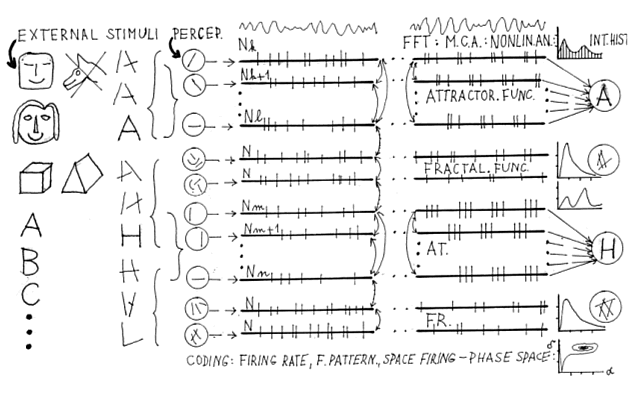
Obr. 7.:Pod vlivem zevní afferentace, učení a "kulturogenese" se vytvářejí v elektrogenesi impulsů neuronální sítě programy v podobě atraktorů.

Obr. 8.:Blokové schema kognitivních funkcí
3. Závěr
Autoorganizační kybernetický model Farleye a Clarka se nabízí jako model centrálního nervového systému, protože obsahuje čtyři důležité části mozku = generátor signálů, komplex elementů, diskriminační jednotku a formátor. Bez násilné představy lze vytvořit analogii s mozkovými strukturami. Generátor signálů připomíná největší podkorové jádro thalamus, kterým také prochází všechny vzruchy ze smyslových orgánů do kortexu a navíc tyto impulsy dostávají rytmus. Kortex připomíná komplex zpočátku neorganisovaných elementů.
Diskrimnační jednotka je velmi podobná limbickému systému v mozku, který také sleduje stavovské veličiny vnitřního i zevního prostředí. Konečně poslední člen soustavy je formátor, který je podobný kmenovým humoroergním modulačním jádrům organisujícím stavy mozkového systému jako je vigilita, synchroní a paraodxní spánek a za patologickýchh okolností i epileptický paroxysmus. Tato jádra kmene a také epileptický fokus nazýváme formátory.
4. Literatura
BENEŠ J.: The cybernetic systems with automatic organisation. Academia. Praha, 1966, p. 174.
COHEN D.B.: Eye movements during REM sleep: The influence of personality and presleep conditions. J.of Personality and Soc. Psychology. 1975, 32, p. 1090-1093.
CRANDALL P.H., DYMOND A., VERZEANO M.: Neural activity in patients with psychomotor epilepsy. Electroenceph.clin.Neurophysiol.,1972, 32, p.98-101
CREUTZFELDT O., OJEMAN G., LETTICH E.: Neuronal activity in the human lateral temporal lobe. Exp. Brain Res. 1989, 77, p. 451-475
FABER J.: Vigilance, sleep, petit mal and EEG as manifestations of programmed brain regulation. Acta Univ. Carol. Med., Monogr. LXXXVII, Praha 1978.
FABER J., VLADYKA V.: Antiepileptic effects of electric stimulation of the locus caeruleus in man. Activ. Nerv. Super. (Praha) 1983, 25, p. 304-308.
FABER J., VLADYKA V.: Nocturnal sleep stereo-EEG and polygraphy in epileptics. Acta Univ. Carol. Med., Monogr.CVIII, Praha 1984.
FABER J., VLADYKA V.: Epileptogenesis and "Psychosogenesis", antithesis or synthesis? Acta Univ. Carol. Med. 1987, 33, p. 245-312.
FABER J.: Temporal epilepsy and consciousness. Triton, Praha 1998,pp. 212, (in Czech). FOERSTER H. VON: On self-organizing systems and their environments. In: Self-organizing systems. M. C. Yovits, S. Cameron (Eds.), Pergamon Press, London 1960.
HEBB D. O.: Textbook of psychology. Philadelphia, Saunders 1972.
JOUVET M., DELORME F.: Locus caeruleus et sommeil paradoxal. Soc. Biol. 1965, 159, p. 895-902.
MADSEN K. B.: Motivational theory. Academia, Praha 1972 (in Czech).
MORUZZI G., MAGOUN H.W.: Brain stem reticular formation and activation of the EEG. Electroenceph.clin. Neurophysiol. 1949, 1, p.455-473.
NICOLIS J. S., PROTONOTARIOS E., LIANOS E.: Some view on the role of noise in "self" - organizing systems. Biol. Cybernetics 1975, 17, p. 183-193.
PETROVICKÝ P., DRUGA R.: The small neuroanatomy. SPN, Charles University, Praha 1989 (in Czech).
SERVÍT Z.: Jan Marek Marci of Kronland. Veda, Slovenská akadémia vied, Bratislava 1989 (in Czech).
SJOESTROEM R.: Effects of psychotherapy in schizophrenia. Acta psycgiatr. scand. 1985, 71, p. 513-522.
TRIMBLE M R.: Interictal Psychoses of Epilepsy. Acta Psychiat. Scand. 1984, Suppl. No.313,vol.69,p. 9-20.